
项目文章 | 重磅NG!中科院昆明植物所李德铢研究员团队揭示竹类植物生活史转变和物种多样化遗传机制
多倍化(基因组加倍)是进化的关键力量,然而,在亚基因组水平上,对于多倍化如何促进植物适应性演化和物种多样性形成的认识仍然不足。竹类植物属于禾本科竹亚科(Bambusoideae),包括二倍体草本竹子(126种)和三种多倍体木本竹子(1576种),其中木本竹类包括温带木本分支、新热带木本分支(均为四倍体)和古热带木本分支(六倍体)。先前研究表明:木本竹子中大约在20百万年前发生的两次独立四倍体化和一次六倍体化事件。木本竹子的染色体数通常保持稳定(例如,四倍体的2n=(40)46-48,六倍体的2n=70-72),这表明组成亚基因组可能一直保持不重排。因此,竹子为研究古老多倍体植物中亚基因组优势的演化提供了一个理想的模型。

2024年3月15日,中国科学院昆明植物研究所李德铢研究员团队在国际顶级期刊《Nature Genetics》(IF=30.8)上在线发表了题为“Genome assemblies of 11 bamboo species highlight diversification induced by dynamic subgenome dominance”的研究论文。该研究选择竹属(禾本科:竹亚科)内11种具有不同倍性的竹子样本,涵盖二倍体(草本植物)到四倍体和六倍体(木本植物),将其从头组装至染色体水平获得这11种竹子的高质量参考基因组,结合比较基因组学、转录组学以及甲基化等进行联合分析,发现木本竹子的亚基因组具有惊人的核型稳定性,两个四倍体谱系中表现出平行的亚基因组优势,并在六倍体谱系中逐渐转移优势。异源多倍体化和亚基因组优势塑造木本竹子的木质茎,快速生长和同步开花等特征的演化。此项工作提供了对多倍体系统中基因组优势的见解,包括其依赖于基因组环境以及其能够随着演化时间改变哪些亚基因组处于优势地位。希望组为本研究提供部分ONT和Hi-C基因组测序服务。
该研究选择了11种具有代表性的竹子为研究对象,包括2个草本竹子(HBs、2x)和9个木本竹子(WBs),其中木本竹子包括3个分支:温带(TWBs、4x)、新热带(NxWBs、4x)和古热带(PWBs、6x),利用ONT+Hi-C的策略从头组装了这11个物种的高质量参考基因组,基因组大小平均分别为625.9 Mb(二倍体)、1,628.3 Mb(四倍体)和1,122.4 Mb(六倍体),Contig N50的平均值和最大值分别为5.3 Mb和17.5 Mb。二代数据的比对率、LTR组装指数(LAI>=10)和BUSCO值(93.2~99.0%)等评估均支持了这些基因组组装的高连续性和完整性。 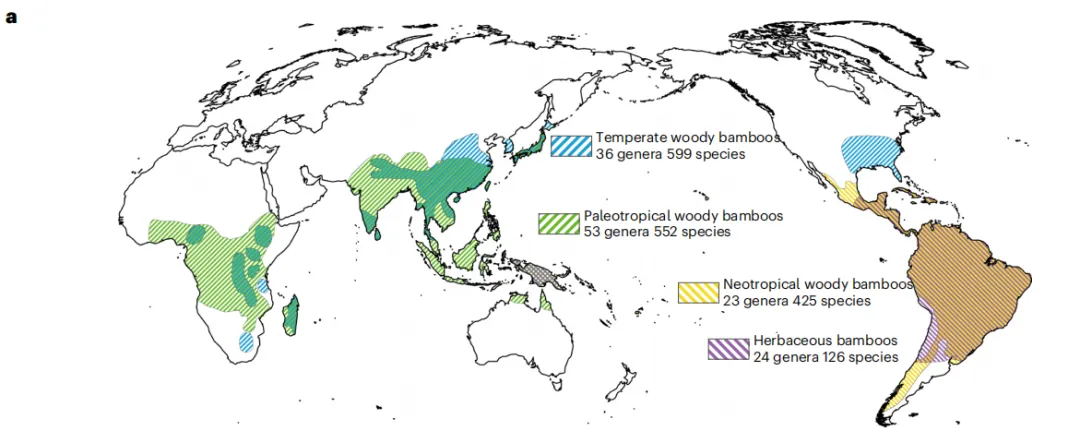
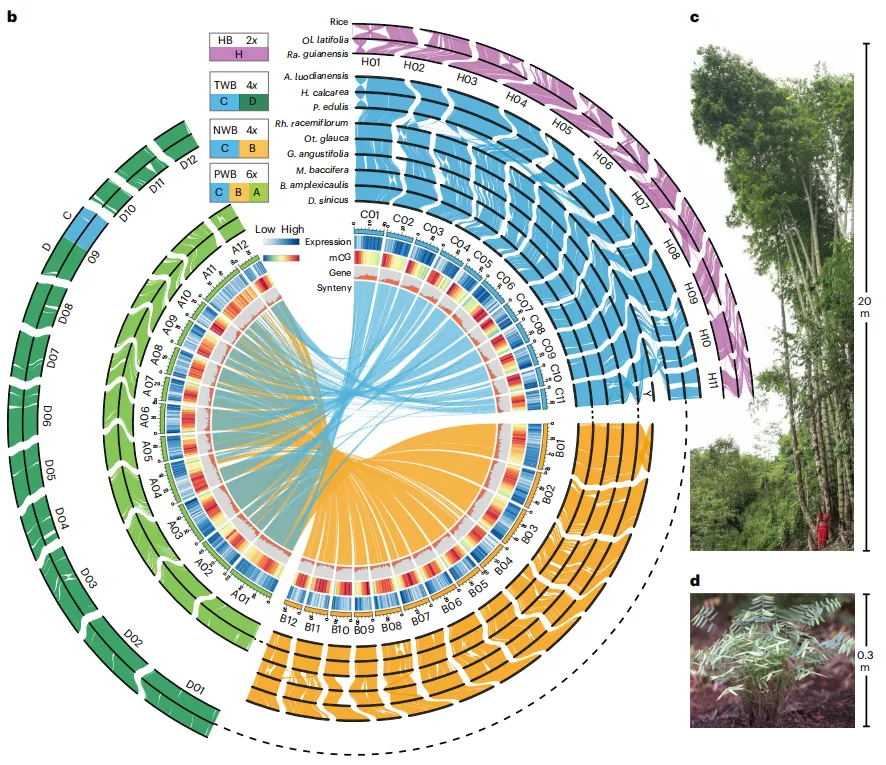
图1 11种竹子基因组特征和共线性图谱
基于系统发育和序列相似性来确定竹子的亚基因组,研究组建了两个同源基因数据集(456个“完美拷贝”和13891个“低拷贝”同源基因)用于系统发育分析,WB中的A、B、C、D和HB中的H亚基因组均能在两个数据集中的到验证。由于基因树之间存在广泛的拓扑结构不一致,研究推断系统发育网络和假定的渗入事件,推断存在杂交事件,如B和C祖先之间的杂交导致A和D祖先的形成,同时还表明了H和A祖先之间的杂交。古老的HBs 和WBs 祖先之间也发生过杂交。总而言之:该研究提出了竹类起源和多倍化的模型,包括不同时间点的分化和多倍化事件,最终形成了不同的竹类群体。
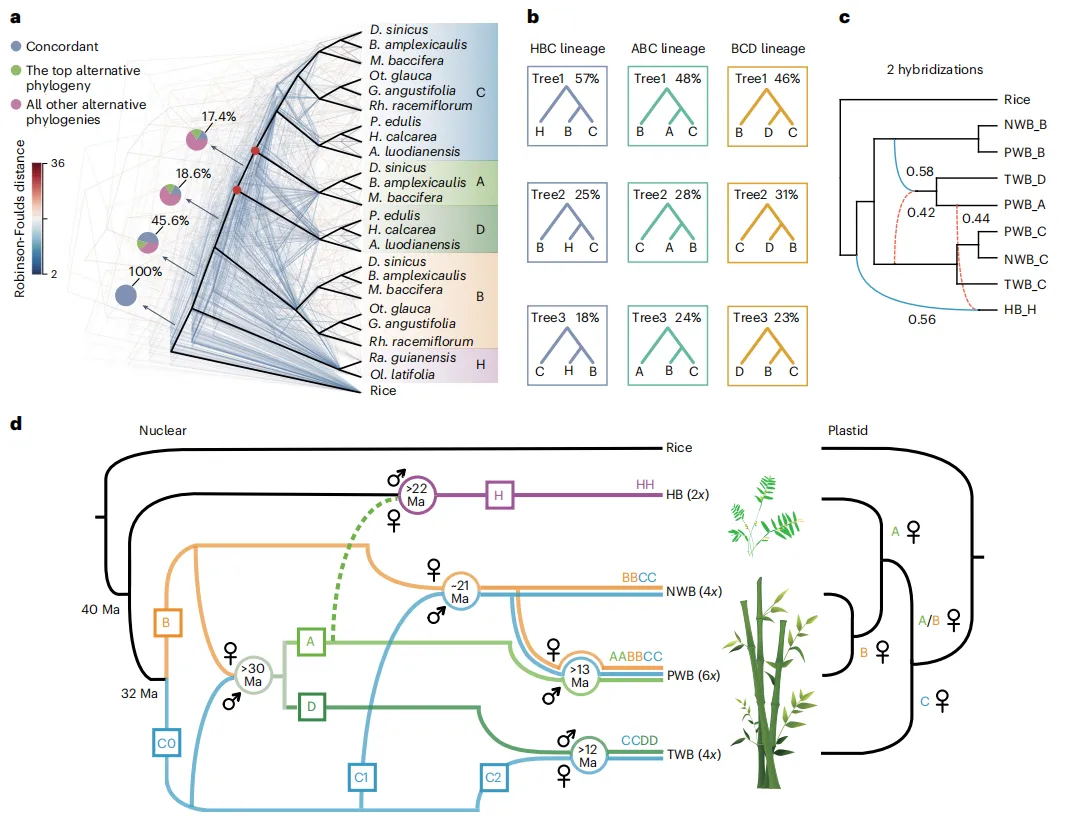
图2 竹类植物主要谱系大的起源与演化
木本竹类展现出非常保守的核型演化特征,尤其是A、B和D亚基因组维持了禾本科祖先的12条染色体状态。大多数裂变和融合事件发生在H和C亚基因组中,HB中的这些事件在很大程度上具有物种特异性,但在C亚基因组中的许多基因分别由热带和温带分支中的不同物种共享,这表示多倍体化在诱导基因组重排中可能起了作用。从亚基因组大小、重复基因丢失、转座子和甲基化等方面研究各个亚基因组间的差异,研究发现:C亚基因组在WB的四个亚基因组中展现出显著不同,其大小与D亚基因组接近,但较小于A和B亚基因组,与TE含量密切相关。多倍体化后,亚基因组和多倍体化后的四六倍体之间存在TE动态多样性。基因进化受多倍体化影响,WBs中观察到全基因组复制丧失。上述分析结果证明:两个四倍体分支中C亚基因组占主导,六倍体中A亚基因组的加入改变了这种主导优势。
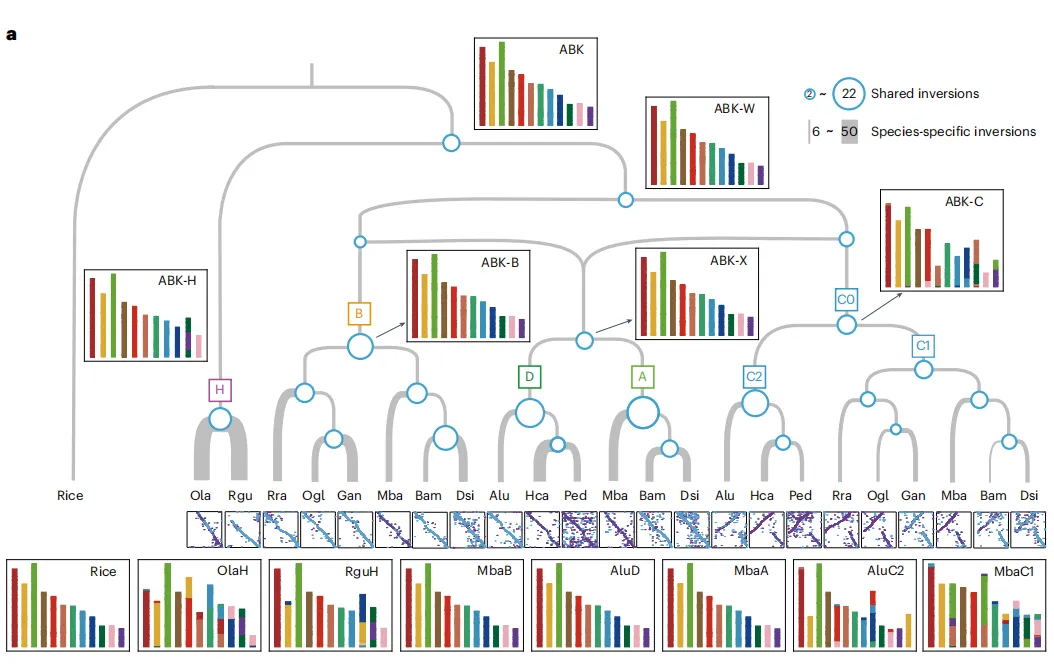
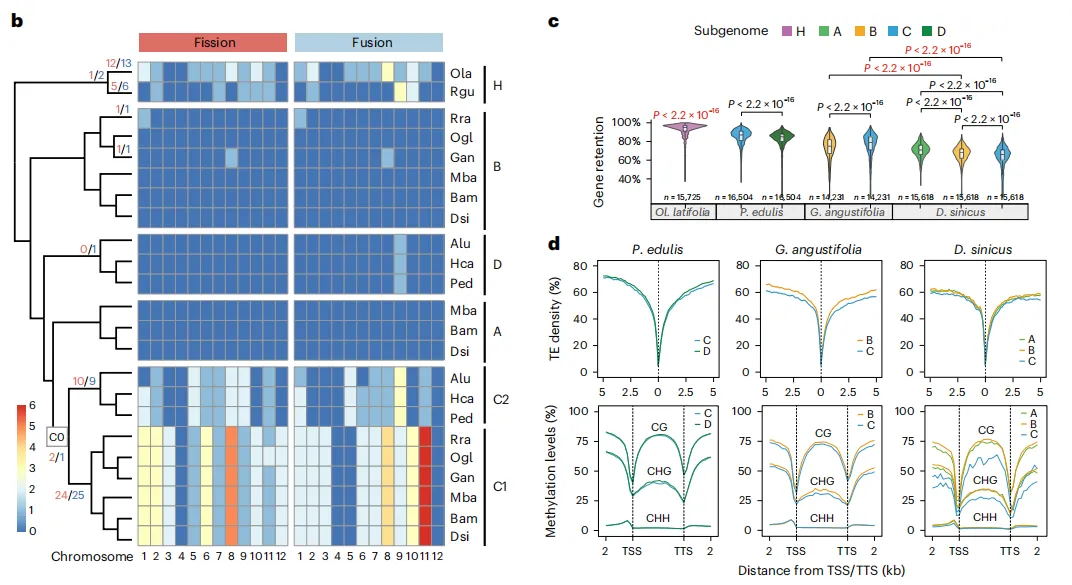
图3 竹类基因组的结构特征与进化
为研究多倍体化后转录图谱的变化,对11个竹子的不同组织在各个发育阶段的476个转录组样本进行测序分析,发现在WBs中的基因其表达丰度较低,与WBs中其他三个亚基因组相比,C亚基因组具有更高比例的表达基因和最高平均表达水平。主成分分析显示跨组织和物种的同源基因之间呈现出一致的基因表达上调和下调模式,而在六倍体(PWBs)中差异较大。同源基因进一步聚类为不同组别,结果显示C亚基因组比D或B亚基因组存在更多的上调基因。基于WGCNA分析探究有偏向性的亚基因组的基因是否更有可能存在共表达,结果显示在四倍体中C亚基因组有更多的共表达基因和hub 基因,而六倍体中A亚基因组拥有更多hub 基因,表明在不同类群中亚基因组的表达模式和共表达关系存在差异。上述结果进一步支持C亚基因组在四倍体分支演化过程中可能具有优势。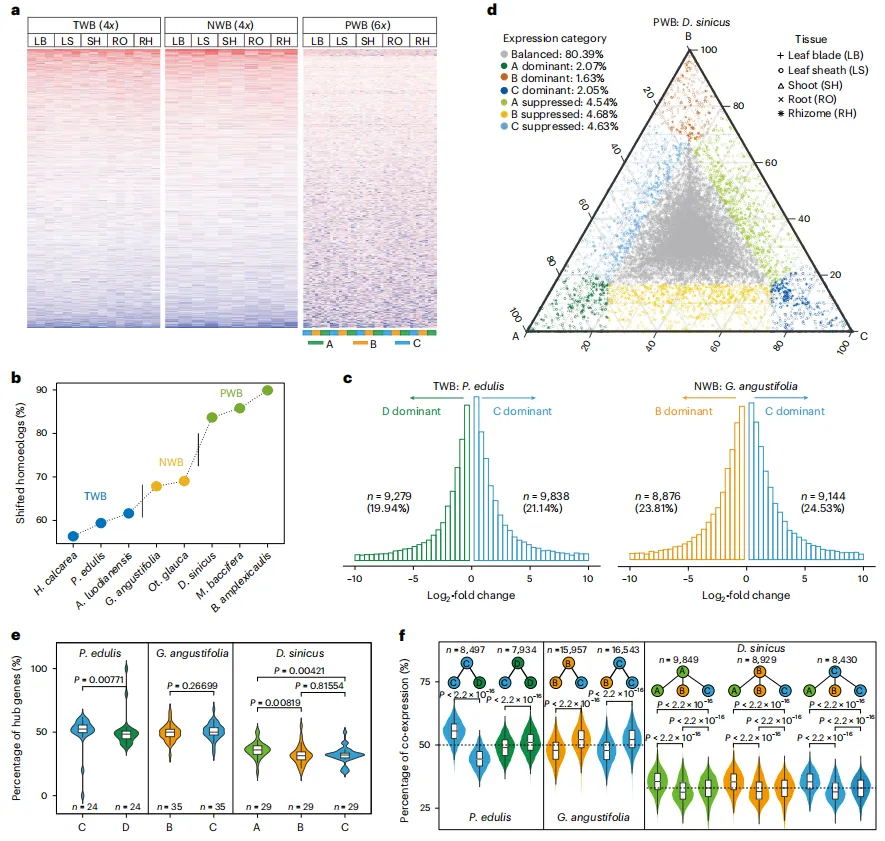
图4 多倍体竹子中的同源基因表达模式
在禾本科中,WBs演化出了包括木质化茎秆和不频繁开花在内的独特性状。为揭示WBs独特性状起源的基因基础,在其演化过程中研究了基因家族大小、新基因和在其演化过程中受到正向选择的基因(PSGs)。三个多倍体谱系中共享了183个PSGs家族,其中C亚基因组的PSG富集。此外,经历两次或更多基因组变化的基因在C亚基因组中占有更高比例。其中许多基因可能参与竹子独特的生命周期,如调控开花的关键因子GI和SPL7,均来自C亚基因组。
功能富集分析表明,在全基因组和亚基因组水平上,特别是C亚基因组的扩张基因家族主要与植物营养生长和发育有关(例如,“植物激素信号转导”和“苯丙素生物合成”)。
进一步研究竹子中木质素生物合成途径中的基因组变化,结果发现,相对于HBs,WBs中几乎所有的木质素相关基因都通过多倍化产生出多拷贝,且COMT 和F5H1 出现串联重复。该途径中检测到有31个基因,其中大多数经历了基因组变化,其中重点关注的COMT,在巨型竹子D. sinicus 的木质化发挥关键作用,其主要负责S型单木质素的生物合成,对与禾本科植物的茎杆强度至关重要。
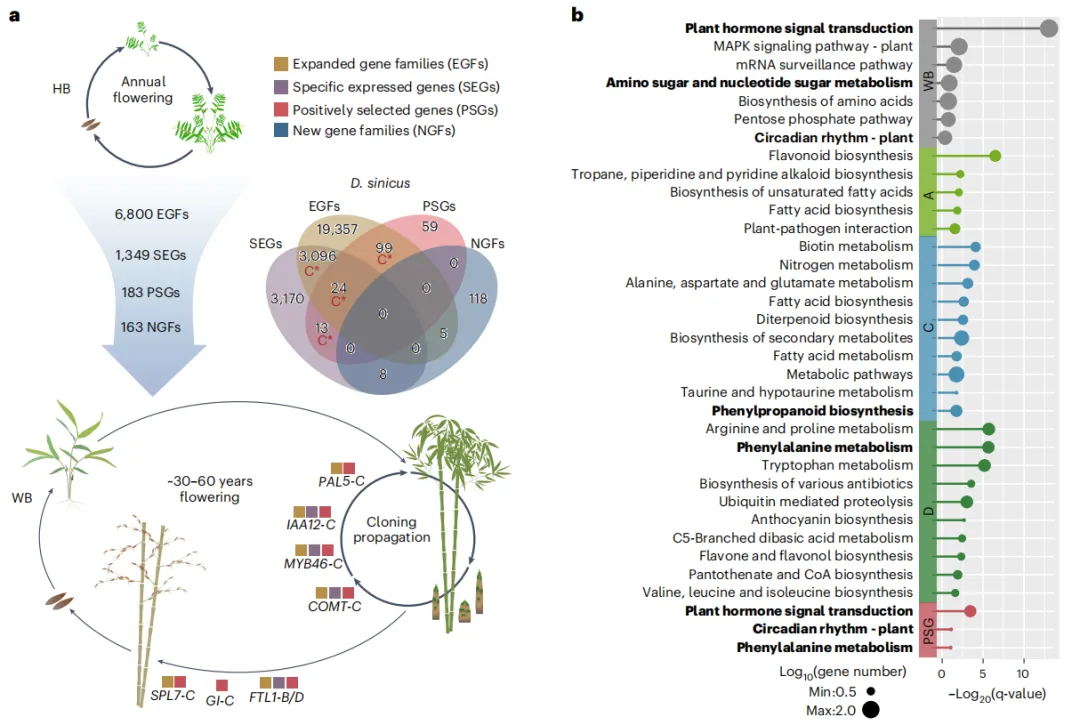
图5 WBs演化的基因组基础
该研究首次提出在多倍体植物类群中可能存在亚基因组优势转移现象,为研究竹类植物物种多样化和独特性状的起源提供了多组学证据,也为深入理解多倍化在植物演化中的重要作用提供了一个难得的案例。




发表评论
想参加讨论吗?请尽情讨论吧!